Мембранный потенциал нейрона. Перенос веществ через мембрану. Мембранный потенциал нейрона Механизм синаптической передачи
Нервная система
регулирует деятельность
организма благодаря
проведению информации
(возбуждения) по сети
нервных клеток.
Цель нейрофизиологии –
это понять биологические
механизмы, которые лежат
в основе проведения
информации по нервной
системе.
Нейроны проводят информацию на большие
расстояния с помощью электрических сигналов,
которые распространяются по аксону.
Используется специальный тип электрических
сигналов – нервный импульс или потенциал
действия.
Потенциал действия является основным
носителем информации в нервной системе
Мембранный потенциал покоя нейрона
Процесс генерации и распространения ПДпроисходит на мембране нейрона.
Клетки, которые способны генерировать и проводить
нервный импульс, имеют возбудимую мембрану.
Мембранный потенциал покоя нейрона
Если на нейрон не действует раздражитель, то оннаходится в состоянии покоя.
В состоянии покоя внешняя сторона мембраны
нейрона заряжена положительно, а внутренняя –
отрицательно. Это состояние называется
мембранным потенциалом покоя.
Мембранный потенциал покоя (МПП) – это
разность потенциалов на мембране нейрона, которую
нейрон имеет в состоянии относительного
физиологического покоя.
Мембранный потенциал покоя нейрона
Потенциал действия – это кратковременноеизменение мембранного потенциала, при котором
внешняя сторона мембраны на одну тысячную
секунду становится отрицательной, а внутренняя –
положительной.
Мембранный потенциал покоя нейрона
1.2.
3.
Чтобы понять, как нейрон передает информацию,
необходимо изучить:
каким образом в состоянии покоя на мембране
нейрона возникает и поддерживается мембранный
потенциал покоя;
каким образом мембранный потенциал
кратковременно изменяется во время генерации
нервного импульса;
каким образом нервный импульс распространяется
вдоль мембраны нейрона.
Мембранный потенциал покоя нейрона
Механизм возникновения МППДвижение ионов
МПП возникает в результате движения ионов
(заряженных частиц) через ионные каналы
мембраны клетки.
Ионы – это атомы или молекулы, которые имеют
положительный (катионы) или отрицательный
(анионы) заряд.
Например, K+, Na+, Cl¯, Ca2+ и т.д.
Механизм возникновения МПП
Движение ионов черезионные каналы связано с
действием двух факторов:
1. диффузия
2. электрическая сила
Диффузия – это движение
ионов из мест c высокой
концентрацией в места с
низкой концентрацией.
Механизм возникновения МПП
ТерминыГрадиент концентрации – это разность
концентрации ионов.
Сила концентрационного градиента – это сила
химической природы, которая перемещает ионы из
мест с высокой концентрацией в места с низкой
концентрацией данного иона.
Правило: чем больше градиент концентрации, тем
больше сила концентрационного градиента.
10. Механизм возникновения МПП
Электрическая сила (I) – этосила, которая перемещает
ионы в электрическом поле.
Электрическая сила
перемещает отрицательные
ионы (анионы) к
положительному заряду
(аноду), а положительные ионы
(катионы) – к отрицательному
заряду (катоду).
11. Механизм возникновения МПП
Движение электрических зарядов в электрическомполе называется электрическим током.
Сила электрического тока определяется двумя
факторами:
1. электрическим потенциалом
2. электрической проводимостью
12. Механизм возникновения МПП
Электрический потенциал (V) – этосила, которая отражает различия в
заряде между катодом и анодом.
Чем больше различия в заряде, тем
больше электрический потенциал, тем
сильнее ток ионов.
Электрический потенциал измеряется
в Вольтах (V).
Электрическая проводимость – это
относительная способность
электрических зарядов двигаться в
электрическом поле.
Чем выше электрическая
проводимость, тем сильнее ток ионов.
13. Механизм возникновения МПП
Электрическое сопротивление (R) – сила,препятствующая движению электрических зарядов.
Электрическое сопротивление измеряется в Омах
(Ω) .
Соотношение между электрическим потенциалом,
сопротивлением и силой тока описывается законом
Ома.
I = V/R
Сила тока равна нулю в двух случаях:
1. либо электрический потенциал равен нулю,
2. либо существует очень большое сопротивление.
14. Механизм возникновения МПП
Движение специфических ионовчерез мембрану под действием
электрической силы может быть
только при одновременном
соблюдении двух условий:
1. мембрана содержит каналы, которые
проницаемы для данного вида ионов;
2. существует разность потенциалов по
обе стороны мембраны.
15. Ионный механизм мембранного потенциала покоя
Мембранный потенциал(МП) – это разность
потенциалов на мембране
нейрона, которую нейрон
имеет в данный момент
времени (Vm).
Мембранный потенциал
нейрона может быть
измерен с помощью
микроэлектрода,
помещенного в цитоплазму
нейрона и подсоединенного
к вольтметру.
16. Ионный механизм мембранного потенциала покоя
В состоянии покоя внутренняя сторона мембранызаряжена отрицательно, а внешняя сторона –
положительно.
Мембранный потенциал покоя (МПП) типичного
нейрона примерно равен - 65 mV.
Vm = -65 mV
Чтобы понять, каким образом возникает и
поддерживается МПП, необходимо рассмотреть
распределение некоторых ионов внутри нейрона и
окружающей его внешней среде.
17. Ионный механизм мембранного потенциала покоя
Равновесный потенциалРассмотрим гипотетическую клетку при
следующих условиях:
1. внутри клетки концентрация катионов K+ и
анионов А¯ выше, чем во внешней среде,
2. мембрана клетки не содержит ионных
каналов.
В этих условиях, несмотря на наличие
разности концентраций ионов,
1. не будет наблюдаться ток ионов через
мембрану;
2. мембранный потенциал будет равен
нулю.
18. Ионный механизм мембранного потенциала покоя
Ситуация изменится, если в мембране появятсяионные каналы, проницаемые для ионов K+, но
не проницаемые для анионов А¯.
Ионы K+ по градиенту концентрации начнут
перемещаться из клетки во внеклеточную среду.
За счет отрицательных ионов А¯ на внутренней
стороне мембраны начинает скапливаться
отрицательный заряд, а на внешней стороне
мембраны начинает появляться положительный
заряд.
Таким образом, на мембране нейрона начинает
появляться разность потенциалов.
19. Ионный механизм мембранного потенциала покоя
По мере увеличения разности потенциаловначинает возрастать электрическая сила,
которая толкает ионы K+ обратно в клетку (так
как положительно заряженные ионы K+
притягиваются к отрицательно зараженному
слою на внутренней стороне мембраны).
Когда на мембране достигается определенное
значение мембранного потенциала
электрическая сила, стремящаяся загнать
ионы K+ внутрь клетки, становится равной
химической силе градиента концентрации,
которая стремится вытолкнуть ионы K+ из
клетки.
Возникает состояние равновесия, при
котором сила электрической природы и сила
химической природы имеют одинаковое
значение, но направлены в разные стороны, а
движение ионов K+ приостанавливается.
20. Ионный механизм мембранного потенциала покоя
Ионный равновесный потенциал – это разностьпотенциалов на мембране, при которой сила химической и
электрической природы уравновешивают друг друга по
отношению к данному иону.
Например, калиевый равновесный потенциал равен
примерно – 80 mV.
Вывод: появление мембранного потенциала в нейроне
происходит автоматически при соблюдении двух условий:
1. существует разница концентраций ионов между внешней и
внутренней средой нейрона;
2. существует избирательная проницаемость мембраны
нейрона для данного иона.
21. Ионный механизм мембранного потенциала покоя
22. Ионный механизм мембранного потенциала покоя
Разница концентраций различных ионов вреальном нейроне
В реальном нейроне разные ионы по разному
распределены во внутриклеточной и внеклеточной
среде.
Ионы
Внеклеточная
концентрация
Внутриклеточная
концентрация
Отношение
Равновесный
потенциал
K+
5
100
1:20
-80 mV
Na+
150
15
10:1
62 mV
Ca2+
2
0,0002
10000:1
123 mV
Cl¯
150
13
11,5:1
-65 mV
23. Ионный механизм мембранного потенциала покоя
Каждый ион имеет свой собственныйравновесный потенциал.
Правило – концентрация ионов K+ больше
внутри клетки, а ионов Na+ и Cl¯ во
внешней среде.
Разница концентраций различных ионов
возникает в результате работы нескольких
ионных насосов, которые встроены в
мембрану нейрона.
24. Ионный механизм мембранного потенциала покоя
Два ионных насоса особенно важныдля понимания работы нейрона:
1. натрий-калиевый
2. кальциевый насос
Натрий-калиевый насос,
используя энергию АТФ, выкачивает
из клетки ионы Na+ и закачивает в
клетку ионы K+ против градиента
концентрации этих ионов.
За один цикл насос выкачивает
3
иона Na+ и 2 иона K+.
На работу этого насоса тратится
больше 70% всей АТФ,
находящейся в мозге.
25. Ионный механизм мембранного потенциала покоя
Кальциевый насос выкачивает из нейрона ионы Ca2+против градиента его концентрации.
1.
2.
Кроме того существуют дополнительные механизмы,
которые обеспечивают уменьшение концентрации ионов
Ca2+ в цитоплазме нейрона (0,00002 mM):
внутриклеточные белки, которые связывают данные
ионы;
клеточные органеллы (в частности, митохондрии и
эндоплазматический ретикулум), которые депонируют
(изолируют) ионы Ca2+.
26. Ионный механизм мембранного потенциала покоя
Значение ионных насосовБез ионных насосов в нейроне не смогла бы
поддерживаться разность концентрации
различных ионов, а, следовательно, в
нейроне не мог бы существовать
мембранный потенциал покоя, без которого, в
свою очередь, нейрон бы не смог отвечать на
внешнее воздействие и передавать
возбуждение.
27. Ионный механизм мембранного потенциала покоя
Относительная проницаемость мембраны для разных ионовВ реальном нейроне мембрана нейрона проницаема не для одного, а
для разных ионов.
Однако проницаемость мембраны для разных ионов разная.
Рассмотрим несколько сценариев для ионов Na+ и K+:
1. Если мембрана проницаема только для иона K+, то мембранный
потенциал будет равен калиевому равновесному потенциалу
(примерно -80 mV).
2. Если мембрана проницаема только для иона Na+, то мембранный
потенциал будет равен натриевому равновесному потенциалу
(примерно 62 mV).
3. Если мембрана имеет одинаковую проницаемость для ионов Na+ и K+, то
мембранный потенциал будет равен среднему значению между
натриевым и калиевым равновесным потенциалом (примерно - 9 mV).
28. Ионный механизм мембранного потенциала покоя
4. Если проницаемость мембраны в 40 раз больше для ионов K+, чемдля ионов Na+, то значение итогового мембранного потенциала
опять будет между натриевым и калиевым равновесным
потенциалом, но при этом ближе к калиевому равновесному
потенциалу.
Последний сценарий наиболее близок к ситуации в реальном
нейроне, в котором мембранный потенциал покоя равен -65 mV.
В реальном нейроне в состоянии покоя мембрана имеет высокую
проницаемость для ионов K+ и относительно низкую для ионов Na+.
29. Ионный механизм мембранного потенциала покоя
Вывод: высокая проницаемость мембранынейрона для ионов K+ является основным
источником мембранного потенциала
покоя (МПП), при этом относительная низкая
проницаемость мембраны для других ионов
(особенно ионов Na+) также вносит
определенный вклад в итоговое значение
МПП нейрона.
30. Ионный механизм мембранного потенциала покоя
Регуляция концентрации ионов K+ во внеклеточнойсреде
Мембранный потенциал очень чувствителен к
изменению концентрации ионов K+ во внеклеточной
среде. Например, если концентрация ионов K+ во
внешней среде уменьшится в 10 раз, то мембранный
потенциал покоя изменится от -65 до -17 mV.
Чувствительность мембранного потенциала к
концентрации ионов K+ привела в эволюции к
появлению механизмов, которые тонко регулируют
содержание этих ионов во внеклеточной среде:
1. гематоэнцефалический барьер
2. глиальные клетки (астроциты)
31. Ионный механизм мембранного потенциала покоя
Гематоэнцефалический барьер (ГЭБ) – этомеханизм, обеспечивающий ограниченный доступ
веществ, которые поступают через стенки капилляров,
к нейронам и глиальным клеткам внутри мозга.
Одна из функций ГЭБа – ограничение поступления из
крови ионов K+ во внеклеточную среду, окружающую
нейроны.
32. Ионный механизм мембранного потенциала покоя
Астроциты обеспечиваютрегуляцию концентрации
ионов K+ с помощью
калиевых насосов и
калиевых ионных каналов,
встроенных в их мембрану.
Когда внеклеточная
концентрация ионов K+
возрастает, эти ионы начинают
заходить внутрь астроцитов
через калиевые ионные
каналы.
33. Ионный механизм мембранного потенциала покоя
Вход ионов K+ в цитоплазмуастроцита приводит к повышению
локальной внутриклеточной
концентрации этих ионов,
которые начинают
распространяться по системе
разветвленных отростков в
другие части глиальной клетки.
Таким образом, астроциты
обладают глиальным
буферным механизмом,
который поддерживает
концентрацию ионов K+ во
внеклеточной среде на
постоянном уровне.
34. Ионный механизм мембранного потенциала покоя
ЗаключениеМеханизм возникновения МПП
1. Активность натрий-калиевого насоса обеспечивает и
поддерживает высокую концентрацию ионов K+ во
внутриклеточной среде нейрона.
2. Мембрана нейрона в состоянии покоя обладает высокой
проницаемостью для ионов K+, так как имеет многочисленные
калиевые каналы.
3. Движение ионов K+ через мембрану нейрона по градиенту их
концентрации приводит к появлению отрицательного заряда на
внутренней стороне мембраны и положительного заряда на
внешней стороне мембраны.
4. Разница потенциалов на мембране нейрона может
рассматриваться как заряд электрической батарейки, который
постоянно поддерживается за счет ионных насосов,
работающих на основе энергии АТФ.
Нейрон в отличие от других клеток способен возбуждаться. Под возбуждением нейрона понимают генерацию нейроном потенциала действия. Основная роль в возбуждении принадлежит другому типу ионных каналов, при открытии которых ионы натрия устремляются в клетку. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов - натрия и калия - имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией.
Ответим на вопрос: как ионные каналы открываются и закрываются? В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка -70 мВ (отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить
Рис. 2.6. Работа натриевого потенциалзависимого канала.
А - канал открыт; Б - канал инактивирован; В - канал инактивирован и закрыт; 1 - мембрана; 2 - потенциалзависимый канал; 3 - инактивирующая частица.
поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается (рис. 2.6). Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Как только канал открывается, в цитоплазму нейрона устремляются из межклеточной среды ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме. Такое движение ионов является следствием простого физического закона: ионы движутся по концентрационному градиенту. Таким образом, в нейрон поступают ионы натрия, они заряжены положительно. Другими словами, через мембрану будет протекать входящий ток ионов натрия, который будет смещать потенциал мембраны в сторону деполяризации, т. е. уменьшать поляризацию мембраны. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал -70 мВ, тогда абсолютная амплитуда потенциала составит величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот
Рис. 2.7. Схема событий при возбуждении нейрона.
потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран.
Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении - как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис. 2.6); она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию - состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения.
Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность
Рис. 2.8. Потенциал действия клетки желудочка сердца собаки.
в нервных клетках составляет величину около 1 / 1000 с (1 мс). Описанная последовательность событий приведена на рис. 2.7.
Подобные потенциалы действия могут возникать и в других клетках, назначение которых - возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 2.8). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т. е. натриевая инактивация затянута.
Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению.
Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли.
Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая - во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона.
Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка -70мВ (отрицательность β цитоплазме), а β состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия - около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1 / 1000 с).
Соединение в живых существах органической материи и металлов, использование живой клетки в качестве проводника электрического тока не могут не удивлять. Чем больше подробностей о своем теле я узнаю, тем чаще возникает удивление от простых его действий. Сгибание руки может восприниматься не как привычный, почти не замечаемый процесс, а как результат слаженной работы мышц и сухожилий, получивших команду от мозга, переданную по нервным волокнам с помощью электричества. Разглядывая полосу глубоко переливающейся синевы, возникающей на куске полированного лабрадорита, который я верчу в руках, я воспринимаю излучаемые им электрические волны рецепторами глаз. В рецепторах эта информация кодируется в виде череды электрических импульсов. Она передается в головной мозг, обрабатывается им, а результат всего этого — восприятие таинственно-красивого камня, возникающие при этом эмоции. Матрица, короче:)
Вместе с восхищением возникает и желание узнать — как это все происходит, желание вникать все глубже в подробности каждого процесса.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.

Достаточно взглянуть на картинку слева, чтоб увидеть основное отличие потенциала покоя от потенциала действия: когда мембрана находится в состоянии покоя, на всей ее протяженности снаружи существует положительный электрический заряд, а внутри — отрицательный.

А потенциал действия, проходящий по нейрону от тела клетки до конца аксона, представляет собой изменение внешнего и внутреннего зарядов клеточной мембраны на небольшом участке нейрона: на очень краткий промежуток времени отрицательный и положительный заряды по разным сторонам мембраны меняются местами.
Возникают вопросы: что запускает потенциал действия? Какова его физическая природа, что именно происходит в клетке при его существовании? Каким образом он распространяется? Почему он идет только в одном направлении? Как клетка восстанавливает исходный потенциал покоя после прохождения по ней волны возбуждения?
Итак, первый вопрос: как возникает потенциал действия? Так как передача сигналов от нейронов-источников к нейрону-получателю, или постсинаптическому нейрону (postsynaptic neuron ), происходит в синапсах, запускающий потенциал действия сигнал стоит искать именно там. Через синапс в нейрон приходят сигнальные вещества, но их поступление не обязательно означает запуск возбуждения нейрона. Если бы нейрон реагировал на очень малые количества сигнальных веществ, это было бы так же неудобно, как иметь супер-чувствительный выключатель, срабатывающий, если на него случайно чихнуть.

Необходимо достаточно сильное воздействие определенных сигнальных веществ, чтобы запустить потенциал действия. В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы — временные изменения зарядов мембраны, вызываемые пришедшими через синапс сигнальными веществами .
Постсинаптические потенциалы сильно отличаются от потенциала действия по своей природе. Если потенциал действия имеет одну строго определенную величину, то величина постсинаптических потенциалов зависит только от вида и количества вызвавших их химических веществ, переданных через синапсы. В то время как потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно отличаются по величине и продолжительности. В одном синапсе нейрона приходящий нервный импульс может вызвать изменение мембранного потенциала на 0,1 мВ, а в другом — на 20 мВ. Кроме того, существуют как вызывающие потенциал действия, так и тормозящие его сигналы.
Тело клетки служит своеобразным калькулятором, простейшим аналоговым компьютером, суммирующим все поступающие в него сигналы в виде возникающих постсинаптических потенциалов. Аналоговые компьютеры устроены очень просто. Например можно построить водяной аналоговый компьютер. Если из одной трубы в банку вливается три килограмма воды, а из другой в ту же банку — пять, то взвесив банку и выяснив, что весы показывают вес, равный восьми килограммам, мы делаем вывод, что 3+5=8. На заре развития компьютерной техники использовались и водяные, и прочие аналоговые вычислительные машины, но наиболее продвинутой была, конечно, машина, которая в точности как и нейроны суммирует электрические токи. Операторы таких ЭВМ занимались вычислениями, перетыкая штекеры из одного гнезда в другое, сливая вместе разные токи.
Так вот в нейроне происходит такое же суммирование электрических токов. В процессе этого суммирования может быть перейден порог возбуждения (excitation threshold ): будет достигнуто определенное значение мембранного потенциала, которое запускает потенциал действия. Обрати внимание на то, что для достижения порога возбуждения мембранной потенциал должен увеличиваться. Если мембранный потенциал покоя нейрона равен -70 мВ, то его порог возбуждения может быть равен -55 мВ. Снижение мембранного потенциала например до -90 мВ приводит к тому, что для преодоления порога возбуждения требуется более сильное воздействие, ведь он остается прежним: -55мВ.

В постсинаптическом нейроне, получающем сигналы от двух возбуждающих и одного тормозящего нейронов, могут существовать различные комбинации сигналов, и далеко не каждая из них приводит к возникновению потенциала действия.
В первой схеме картинки вверху изображена ситуация, когда только от одного возбуждающего нейрона приходит слабый сигнал, которого недостаточно для преодоления порога возбуждения. Этот сигнал повторяется, но к моменту прихода второго, такого же слабого сигнала от того же аксона возбуждение мембраны уже достигает потенциала покоя, и потенциал действия не возникает.
На второй схеме изображен тот же самый слабый сигнал, но приходящий с большей частотой. Именно из-за увеличения частоты сигнала мембрана не успевает вернуться в состояние покоя в промежутке между двумя сигналами. В этом случае сумма остаточного заряда мембраны после первого сигнала и поступающего следом второго сигнала позволяет преодолеть порог возбуждения нейрона. В этом случае у основания аксона, которое называется аксонным холмиком (axon hillock [‘aksɒn ‘hɪlək] ), возникает потенциал действия.
На третьей схеме от обоих возбуждающих нейронов приходят слабые сигналы, и их суммы достаточно для запуска потенциала действия.
Четвертая схема показывает воздействие на постсинаптический нейрон возбуждающего и тормозящего сигнала. При поступлении только возбуждающего сигнала мембранный потенциал незначительно увеличивается, а при поступлении только тормозного сигнала — снижается. Если эти два сигнала приходят одновременно, то они всего лишь уничтожают друг друга, оставляя мембранный потенциал в состоянии, близком к потенциалу покоя. Это напоминает надавливание на одну и ту же дверь с разных сторон: по отдельности каждое воздействие могло бы приоткрыть дверь или наоборот, прикрыть ее, но существующие одновременно, они приводят лишь к небольшому сдвигу ее по направлению действия большей по модулю силы.
Природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект. Нейрон «складывает» все поступившие в него возбуждающие постсинаптические потенциалы, «вычитает» поступившие тормозные постсинаптические потенциалы, и как только будет перейден порог возбуждения, возникает потенциал действия.
Чтобы разобраться, что происходит в клетке во время запуска и прохождения потенциала действия , необходимо перечислить каждого участвующего игрока и его действие, как было сделано при описании возникновения мембранного потенциала покоя в . Итак, действующие лица при создании потенциала действия :

1 . , которые были закрыты при существовании мембранного потенциала покоя, открываются, как только потенциал достигает порога возбуждения. Так как внутри клетки на мембране существует отрицательный заряд, то в этот момент через каналы туда врываются притягиваемые им
2 . положительно заряженные ионы натрия (Na +). Именно они создавали положительный заряд на внешней поверхности мембраны нейрона, и именно в этот момент они переносят его внутрь, оставляя на внешней мембране отрицательный заряд, который создают находящиеся вне клетки отрицательно заряженные ионы хлора.
3 . таким образом ионы хлора (Cl —) создают отрицательный заряд на внешней поверхности мембраны.

Перемещение ионов натрия внутрь клетки приводит к резкому изменению зарядов внутри и снаружи нейрона. Место существующего во время покоя нейрона отрицательного внутреннего заряда мембраны занимает положительный заряд ионов натрия. А снаружи клеточной мембраны после ухода оттуда Na + наконец-то может проявить себя Cl — , чей отрицательный заряд больше не компенсируется положительно заряженными ионами. Таким образом потенциал клеточной мембраны внезапно возрастает с -70 до +40 мВ. Именно это повышение, называемое деполяризацией (depolarization ), изображено резко идущей вверх частью пика на этой картинке. Такое повышение мембранного потенциала является сигналом закрыть потенциал-зависимые натриевые каналы, а также открывает потенциал-зависимые калиевые каналы, о которых раньше мы еще не говорили.
4 . потенциал-зависимые калиевые каналы. Они существуют в нейроне в дополнение к проточным калиевым каналам, которые, как ты наверное помнишь из прошлой главы про потенциал, не имеют ворот и обеспечивают постоянный отток калия из клетки, играя существенную роль в формировании потенциала покоя. Так вот оказывается, что есть две разновидности калиевых каналов, и сейчас мы рассмотрим работу потенциал-зависимого калиевого канала. Открываясь после того, как потенциал действия достиг своего пика, потенциал-зависимые калиевые каналы резко увеличивают отток ионов калия из клетки. Существующий в этот момент внутри нейрона положительный заряд, образовавшийся за счет притока Na + , активно выпихивает положительно заряженные ионы калия (К +) из клетки через оба вида каналов.

Итак, еще раз проследим цепочку событий до этого момента: потенциал достигает порога возбуждения, и этот электрический сигнал приводит к тому, что открылись потенциал-зависимые натриевые каналы, ионы Na + хлынули внутрь, притягиваемые отрицательно заряженными протеинами, произошла деполяризация, то есть разница зарядов снаружи и внутри резко снизилась, и даже более того — отсутствие положительно заряженных ионов натрия привело к возникновению отрицательного заряда там, где только что был положительный — произошла смена полюсов. Действие электрического импульса закончилось, натриевые каналы вновь закрылись, и столпившиеся внутри ионы Na + вытолкнули одноименно заряженные ионы К + через открывшиеся потенциал-зависимые калиевые каналы и через проточные калиевые каналы.
Заряд внутренней поверхности клеточной мембраны начинает стремительно снижаться, на схеме этому соответствует идущая вниз часть пика. Такое снижение заряда называется реполяризацией (repolarization ), то есть разница зарядов вне и внутри нейрона вновь восстановилась, как и было изначально в состоянии покоя. Но восстановилась именно разница зарядов, а не ионный состав, ее создающий. Несмотря на то, что в конце реполяризации снаружи мембраны образуется положительный заряд, равный положительному заряду мембраны в состоянии покоя, до достижения мембраной состояния покоя еще далеко. Ведь этот положительный заряд принесли туда вышедшие из клетки ионы К + , а при потенциале покоя его поддерживают ионы Na + , которые в данный момент заперты внутри клетки, в том самом месте, где ранее в состоянии покоя находились ионы калия.
В конце реполяризации может возникнуть снижение мембранного потенциала даже ниже его значения для покоящегося нейрона, или гиперполяризация (hyperpolarization [ˌhaɪpəpələraɪ’zeɪʃn] ). Через короткий промежуток времени она компенсируется дополнительным выходом ионов калия из клетки, и наступает прежнее, стабильное состояние покоя.

Когда мембранный потенциал становится равен потенциалу покоя (-70 мВ), потенциал-зависимые калиевые каналы закрываются. Через проточные калиевые каналы К + уже не вытекает так стремительно, ведь его концентрация внутри клетки упала, а снаружи находится много ионов калия. В то же время внутри находится много ионов натрия, которые должны быть выведены из клетки для того, чтоб через нее мог снова пройти потенциал действия.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na + и одновременно проносящий внутрь два К + . О его работе было рассказано в предыдущей главе про потенциал, и он схематично показан в четвертой части этой картинки. Совместные действия натрий-калиевого насоса и проточных калиевых каналов приводят к тому, что с внешней стороны мембраны опять возникает большая концентрация Na + , а концентрация К + внутри нейрона выше наружной.
Если бы работали только проточные калиевые каналы, то ионы калия только выходили бы из клетки, и было бы невозможным поддерживать внутри большую концентрацию К + , чем снаружи. Если бы действовали только потенциал-зависимые натриевые каналы, то ионы натрия вообще не могли бы покинуть клетку после того, как вошли туда, создавая потенциал действия. Но натрий-калиевый насос (ну то есть натрий-калиевый канал) решает обе эти проблемы: он выводит наружу Na + , восстанавливая его высокую концентрацию снаружи. И загоняет внутрь прошедшие через проточные каналы К + , не давая им возможности распределиться с обеих сторон мембраны равномерно, лишившись таким образом парциального давления, необходимого для формирования мембранного потенциала покоя.
Через тысячную долю секунды после того, как реполяризация нейронной мембраны достигла минимума, активная работа мембранных каналов восстанавливает потенциал покоя. Нейрон готов к проведению следующего потенциала действия.
5 . положительно заряженные ионы кальция (Ca 2+) играют важную роль в формировании потенциала действия лишь в некоторых нервных клетках, поэтому в данной главе, содержащей общую для различных нейронов информацию, она не будет описана.

На этом общее описание перемещения ионов через клеточную мембрану, превращающее потенциал покоя нейрона в потенциал действия, закончено. Можно сделать перерыв и переварить его вместе с чем-нибудь вкусным, заодно подпитав мозг, компенсировав те калории, которые он затратил на работу по обработке всей этой информации.
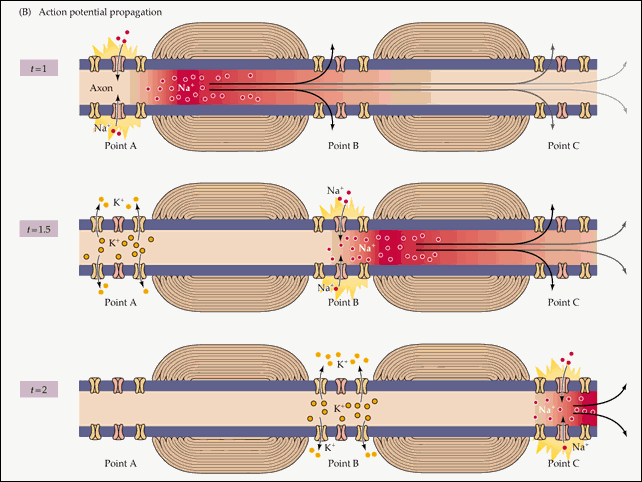
Распространение потенциала действия по всей длине аксона обеспечивается тем фактом, что возникший при его существовании в одном участке мембраны нейрона электрический ток возбуждает соседние участки, открывая натриевые потенциал-зависимые каналы и запуская уже там потенциал действия. Точнее, электрический ток запускает потенциал действия только с одной стороны от места существования предыдущего потенциала. Потому что с другой стороны натриевые каналы инактивированы после недавнего открытия, а без их участия возникновение потенциала действия невозможно. Если бы не было этого временного промежутка нефункциональности между двумя открытиями натриевых каналов, потенциал действия не мог бы распространяться только в одном направлении. Происходящая в результате цепная реакция открытия натриевых каналов обеспечивает передачу нервного импульса от места его возникновения до окончаний аксона, подходящих к другим нейронам (или к мышечной клетке).

Схематично эту цепную реакцию можно изобразить так:
Момент времени 1 : Электрический ток, возникший на в результате открытия потенциал-зависимых натриевых каналов на участке 1 , возбуждает соседние участки мембраны.
Момент времени 2 : В результате этого возбуждения на участке 2 открываются потенциал-зависимые натриевые каналы и возникает потенциал действия, который, в свою очередь, возбуждает соседние участки мембраны. На участке 1 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал после прохождения потенциала действия.
Момент времени 3 : Электрический ток, возникший в результате открытия потенциал-зависимых натриевых каналов на участке 2 , вызвал открытие таких же каналов на участке 3 , и потенциал действия перешел туда. На участке 1 потенциал-зависимые натриевые каналы не смогли открыться, несмотря на электрическое возбуждение от участка 2 , так как они были временно инактивированы после создания потенциала действия. На участке 2 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал покоя после прохождения потенциала действия.
Мы уже знаем, что отростки нейронов могут быть покрыты обернутыми вокруг них клетками нейроглии, или миелиновой оболочкой. Именно она делает аксон похожим на вытянутую цепочку сосисок, про ее образование и функции говорилось в . Передача потенциала возбуждения по «голым», не покрытым миелиновыми клетками, нервным волокнам отличается от его прохождения по миелинизированному аксону. При сравнении заметны существенные преимущества, предоставляемые обернутыми вокруг аксона клетками нейроглии:

1 . Экономичность . Мембрана аксона под миелином не имеет каналов для пропускания ионов, создающих потенциал действия. Каналы существуют только в узких промежутках между глиальными клетками, которые называют перехваты Ранвье (Ranvier’s constriction ), и только на этих узких участках аксона возникает потенциал действия. Концентрация ионных каналов в этих перехватах в 100 раз выше, чем в мембранах безмиелиновых волокон, тем не менее для того, чтобы обеспечить их работу, требуется значительно меньше энергии по сравнению с тем, сколько энергии потребовалось бы для обеспечения ионных каналов, расположенных по всей поверхности «голого» аксона. Кроме того, такая экономичность позволяет быстрее «перезаряжать» аксон для проведения следующего потенциала действия, поэтому миелинизированные волокна способны передавать сигналы более высокой частоты.
2
. Скорость
. По «голому» аксону сигнал передается в каждой его точке. Ионные каналы открываются в каждой точке его поверхности, один за другим, что напоминает тщательное, скрупулезное вчитывание в текст.
В миелинезированном аксоне потенциал действия, возникший в одном перехвате Ранвье, своим электрическим полем дотягивается до соседних перехватов, что приводит к возникновению в них новых потенциалов действия. То есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает следующий после поврежденного перехват, а в случае необходимости даже третий, четвертый и пятый по счету. Распространение электрического поля на такую длину возможно из-за электроизоляции, создаваемой миелиновыми муфтами: она уменьшает рассеивание электрического поля, которое неизбежно для безмиелиновых волокон.
Таким образом, скорость распространения потенциала действия по миелинизированным волокнам намного выше по сравнению с немиелинизированными. Ведь за один и тот же отрезок времени потенциал действия на «голом» аксоне возбудит лишь находящийся непосредственно рядом участок, а потенциал действия на миелинизированном волокне перепрыгнет на расстояние, равное расстоянию между одним или несколькими перехватами Ранвье:


Диапазон скоростей передачи потенциала действия в различных миелинизированных волокнах очень широк: от нескольких метров в секунду до «мирового рекорда», установленного аксоном креветки: он проводит возбуждение со скоростью, превышающей 200 м/с.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения потенциала действия за счёт миелиновой оболочки, достаточно сравнить скорость его распространения по немиелинизированным и миелинизированным волокнам нейронов человека. Если взять скорость проведения импульса в одинаково тонких волокнах, то в безмиелиновом волокне она составит примерно 1 м/с, а в миелинизированном 15-20 м/с. В толстых аксонах человека, покрытых миелиновой оболочкой, скорость проведения может достигать 120 м/с.
Итак, подведем итог : передача электрического импульса через тело и отростки нейрона осуществляется с помощью прохождения ионов через клеточную мембрану.

1 . сначала на клеточной мембране создается потенциал покоя . Его образуют положительно заряженные ионы натрия (Na + ) снаружи клетки и отрицательно заряженные протеины внутри клетки. Создающие заряды частицы расположены очень близко к клеточной мембране, ведь они имеют разные знаки и притягиваются друг к другу. Поэтому потенциал покоя существует только возле мембраны, в целом среда внутри нейрона и вне его электрически нейтральна. Кроме Na + и отрицательно заряженных протеинов, в создании и поддержании электрического заряда участвуют положительно заряженные ионы (К + ) и отрицательно заряженные ионы хлора (Cl — ) , тоже находящиеся вблизи мембраны. Потенциал покоя равен примерно -70 мВ (в дальнейшем при указывании численного значения мембранных потенциалов слово «примерно» будет опускаться).
2 . когда нейрон через синапсы получает сигналы от других нейронов, под их влиянием на мембране тела нейрона начинают возникать постсинаптические потенциалы , то есть изменения величины электрических зарядов на внутренней и внешней поверхностях мембраны. Под влиянием этих сигналов внутренний отрицательный заряд и внешний положительный могут увеличиваться или уменьшаться на разные величины. И нейрон выступает в роли калькулятора, суммирующего эти воздействия.

3 . итогом суммирования постсинаптических потенциалов может являться продолжение нахождения нейрона в состоянии покоя. Но если сумма дойдет до значения, называемого порогом возбуждения , то в аксонном холмике (утолщении тела нейрона у основания аксона) возникнет потенциал действия .
4 . в создании потенциала действия главную роль играет клеточная мембрана . Она регулирует прохождение через нее ионов (всех перечисленных, кроме отрицательно заряженных протеинов, которые не могут покидать пределы клетки), открывая и закрывая специализированные каналы, способные проводить только определенные ионы.
5 . когда мембрана нейрона переходит порог возбуждения, в ней открываются потенциал-зависимые натриевые каналы . Исчезает то препятствие, которое отделяло положительно заряженные Na + от притягивающих их отрицательно заряженных протеинов, и Na + устремляются внутрь клетки . Таким образом наружный положительный заряд переносится внутрь клетки, а снаружи остается отрицательный заряд, создаваемый ионами Cl — . Происходит изменение мембранного потенциала от потенциала покоя, равного -70 мВ, до +40 мВ, и это называют потенциалом действия . Важно понимать, что это изменение потенциала происходит только на том участке мембраны, где открылись потенциал-зависимые натриевые каналы.
6 . В момент достижения мембраной потенциала в 40 мВ потенциал-зависимые натриевые каналы закрываются, и на этом же участке открываются потенциал-зависимые калиевые каналы . Снаружи в этот момент существует отрицательный заряд, а внутри — положительный, создаваемый вошедшими туда ионами Na + и уже находящимися там в большом количестве ионами К + . Ионы калия выпихиваются в окружающую нейрон среду через открывшиеся каналы, что резко снижает внутренний заряд мембраны и увеличивает внешний. По обеим сторонам мембраны создаются заряды той же величины, которые обычно поддерживают потенциал покоя. Но это состояние клетки отличается от потенциала покоя тем, что ионы калия и натрия находятся не на своих местах: Na + находится внутри, а К + снаружи. Опять же все описанное касается только одного места на мембране, в котором только что перестал существовать потенциал действия.
7 . Чтобы переместить положительно заряженные ионы Na + и К + по нужные для поддержания потенциала покоя стороны мембраны, используется натрий-калиевый насос . И это действие тоже касается ограниченного участка мембраны: того, где только что закрылись потенциал-зависимые калиевые каналы.
Один цикл работы натрий-калиевого насоса заключается в том, что он захватывает изнутри клетки три иона Na + , перемещает их наружу, где захватывает два иона К + и перемещает их внутрь . То есть канал обменивает три иона Na + на два иона К + , причем Na + могут перемещаться только наружу, а К + только внутрь клетки. Таким образом восстанавливается исходная концентрация натрия на наружной стороне мембраны и калия на внутренней. Натрий-калиевый насос выводит из клетки три положительно заряженных иона, а вводит в нее только два. Следовательно, при каждом его цикле внутренний положительный заряд мембраны уменьшается, а внешний увеличивается. К тому же и вытекающие ионы калия уносят изнутри наружу свои положительные заряды. Возникает вопрос: если потенциал мембраны уже и так был равен потенциалу покоя после входа ионов калия внутрь клетки, зачем его снижать еще сильнее? Ответ заключается в том, что кроме упомянутых здесь ионов, существуют и другие, чье участие в формировании потенциала покоя менее существенно, и именно для компенсации их влияния необходимо такое выравнивание мембранного потенциала. Итогом этого процесса является достижение мембранного потенциала покоя: заряд внутренней стороны мембраны отрицателен, внешней положителен, разность потенциалов равна -70 мВ.
8 . Итак, в том месте клеточной мембраны, где всего три шага назад существовал потенциал действия, опять царит потенциал покоя. Но потенциал действия не прошел бесследно: его существование (описанное в пункте 5) создало электрическое поле, возбудившее соседние с ним части мембраны . С одной стороны (по направлению к концу аксона) это возбуждение вызвало открытие потенциал-зависимых натриевых каналов, запустив уже на новом участке потенциал действия. Который в свою очередь порождает электрическое поле, запускающее следующий потенциал действия, и этот цикл повторяется по всей длине аксона, от его основания до концов, просоединенных к следующим в нервной цепи клеткам.
С другой стороны от потенциала действия, ближе к телу нейрона, несмотря на воздействие электрического поля потенциал действия не возникает. Потому что потенциал-зависимые натриевые каналы на этом участке совсем недавно открывались, и на некоторое время они инактивированы. Именно эта пауза в их работе обеспечивает распространение потенциала действия только в одном направлении.
9 . Любой сигнал, будь то передача информации от сенсорного нейрона в мозг, или команда мозга, идущая к мотонейрону, передается одинаковыми по величине потенциалами действия . Результат потенциала действия (появление восприятия запаха, обусловленное работой сенсорных нейронов, или перемещение тела поближе к его источнику в результате слаженной работы мотонейронов) зависит исключительно от того, в какую нервную цепь включен нейрон, этот потенциал передающий. Для передачи интенсивности сигнала (силы запаха или величины сокращения мышцы) используется частота, с которой потенциалы действия передаются по нейрону. Слабый сигнал передается редкими, а сильный — частыми потенциалами действия .
10 . миелиновая оболочка вокруг отростков нейрона повышает скорость распространения потенциала действия за счет того, что сигнал передается на большое расстояние, равное ширине одной обернутой вокруг отростка клетки нейроглии, а не на непосредственно прилегающий к возбужденному участок, как это происходит у безмиелиновых волокон.
- управляемые. По механизму управления: электро-, хемо- и механоуправляемые;
- неуправляемые. Не имеют воротного механизма и всегда открыты, ионы идут постоянно, но медленно.
Потенциал покоя — это разность электрических потенциалов между наружной и внутренней средой клетки.
Механизм формирования потенциалов покоя. Непосредственная причина потенциала покоя — это неодинаковая концентрация анионов и катионов внутри и вне клетки. Во-первых, такое расположение ионов обосновано разницей проницаемости. Во-вторых, ионов калия выходит из клетки значительно больше, чем натрия.
Потенциал действия — это возбуждение клетки, быстрое колебание мембранного потенциала вследствие диффузии ионов в клетку и из клетки.
При действии раздражителя на клетки возбудимой ткани сначала очень быстро активируются и инактивируются натриевые каналы, затем с некоторым опозданием активируются и инактивируются калиевые каналы.
Вследствие этого ионы быстро диффундируют в клетку или из нее согласно электрохимическому градиенту. Это и есть возбуждение. По изменению величин и знака заряда клетки выделяют три фазы:
- 1-я фаза — деполяризация. Уменьшение заряда клетки до нуля. Натрий движется к клетке согласно концентрационному и электрическому градиенту. Условие движения: открыты ворота натриевого канала;
- 2-я фаза — инверсия. Изменение знака заряда на противоположный. Инверсия предполагает две части: восходящую и нисходящую.
Восходящая часть. Натрий продолжает двигаться в клетку согласно концентрационному градиенту, но вопреки электрическому градиенту (он препятствует).
Нисходящая часть. Калий начинает выходить из клетки согласно концентрационному и электрическому градиенту. Открыты ворота калиевого канала;
- 3-я фаза — реполяризация. Калий продолжает выходить из клетки согласно концентрационному, но вопреки электрическому градиенту.
Критерии возбудимости
При развитии потенциала действия происходит изменение возбудимости ткани. Это изменение протекает по фазам. Состояние исходной поляризации мембраны характерно отражает мембранный потенциал покоя, которому соответствует исходное состояние возбудимости а, следовательно, исходное состояние возбудимой клетки. Это нормальный уровень возбудимости. Период предспайка — период самого начала потенциала действия. Возбудимость ткани слегка повышена. Эта фаза возбудимости — первичная экзальтация (первичная супернормальная возбудимость). Во время развития предспайка мембранный потенциал приближается к критическому уровню деполяризации и для достижения этого уровня сила раздражителя может быть меньше пороговой.
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны, и она утрачивает способность отвечать возбуждением на раздражители сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности, т.е. абсолютной невозбудимости, которая длится до конца перезарядки мембраны. Абсолютная рефрактерность мембраны возникает в связи с тем, что натриевые каналы полностью открываются, а затем инактивируются.
После окончания фазы перезарядки возбудимость ее постепенно восстанавливается до исходного уровня — это фаза относительной рефрактерности, т.е. относительной невозбудимости. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Поскольку в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых каналов.
Следующему периоду соответствует повышенный уровень возбудимости: фаза вторичной экзальтации или вторичной супернормальной возбудимости. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, по сравнению с состоянием покоя исходной поляризации, то порог раздражения снижен, т.е. возбудимость клетки повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. Мембранный потенциал увеличивается — возникает состояние гиперполяризации мембраны. Удаляясь от критического уровня деполяризации, порог раздражения слегка повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины.
Механизм возникновения мембранного потенциала покоя
Каждая клетка в состоянии покоя характеризуется наличием трансмембранной разности потенциалов (потенциала покоя). Обычно разность зарядов между внутренней и внешней поверхностями мембран составляет от -80 до -100 мВ и может быть измерена с помощью наружного и внутриклеточного микроэлектродов (рис. 1).
Разность потенциалов между наружной и внутренней сторонами мембраны клетки в состоянии ее покоя называют мембранным потенциалом (потенциалом покоя).
Создание потенциала покоя обеспечивается двумя основными процессами — неравномерным распределением неорганических ионов между внутри- и внеклеточным пространством и неодинаковой проницаемостью для них клеточной мембраны. Анализ химического состав вне- и внутриклеточной жидкости свидетельствует о крайне неравномерном распределении ионов (табл. 1).
В состоянии покоя внутри клетки много анионов органических кислот и ионов К+, концентрация которых в 30 раз больше, чем снаружи; ионов Na+, наоборот, снаружи клетки в 10 раз больше, чем внутри; СI- также больше снаружи.
В покое мембрана нервных клеток наиболее проницаема для К+, менее — для СI- и очень мало проницаема для Na+/ Проницаемость мембраны нервного волокна для Na+ B покое в 100 раз меньше, чем для K+. Для многих анионов органических кислот мембрана в покое совсем непроницаема.
Рис. 1. Измерение потенциала покоя мышечного волокна (А) с помощью внутриклеточного микроэлектрода: М — микрозлектрод; И — индифферентный электрод. Луч на экране осциллографа (В) показывает, что до прокола мембраны микроэлектродом разность потенциалов между М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена отрицательно по отношению к ее наружной поверхности (по Б.И. Ходорову)
Таблица. Внутри- и внеклеточные концентрации ионов мышечной клетки теплокровного животного, ммоль/л (по Дж. Дудел)
|
Внутриклеточная концентрация |
Внеклеточная концентрация |
|
|
А- (анионы органических соединений) |
В силу градиента концентраций К+ выходит на наружную поверхность клетки, вынося свой положительный заряд. Высокомолекулярные анионы не могут следовать за К+ из-за непроницаемости для них мембраны. Ион Na+ также не может возместить ушедшие ионы калия, ибо проницаемость мембраны для него значительно меньше. СI- по градиенту концентраций может перемешаться только внутрь клетки, увеличивая тем самым отрицательный заряд внутренней поверхности мембраны. Вследствие такого перемещения ионов возникает поляризация мембраны, когда наружная ее поверхность заряжается положительно, а внутренняя — отрицательно.
Электрическое поле, которое создастся на мембране, активно вмешивается в распределение ионов между внутренним и наружным содержимым клетки. По мере возрастания положительного заряда на наружной поверхности клетки иону К+ как положительно заряженному становится все труднее перемещаться изнутри наружу. Он движется как бы в гору. Чем больше величина положительного заряда на наружной поверхности, тем меньшее количество ионов К+ может выходить на поверхность клетки. При определенной величине потенциала на мембране количество ионов К+, пересекающих мембрану в том и другом направлении, оказывается равным, т.е. концентрационный градиент калия уравновешивается имеющимся на мембране потенциалом. Потенциал, при котором диффузионный поток ионов становится равным потоку одноименных ионов, идущих в обратном направлении, называют потенциалом равновесия для данного иона. Для ионов К+ потенциал равновесия равен -90 мВ. В миелинизированных нервных волокнах величина потенциала равновесия для ионов СI- близка к значению мембранного потенциала покоя (-70 мВ). Поэтому, несмотря на то что концентрация ионов СI- снаружи волокна больше, чем внутри его, не отмечается их одностороннего тока в соответствии с градиентом концентраций. В этом случае разность концентраций сбалансирована потенциалом, имеющимся на мембране.
Ион Na+ по градиенту концентраций должен был бы входить внутрь клетки (его потенциал равновесия составляет +60 мВ), и наличие отрицательного заряда внутри клетки не должно было бы препятствовать этому потоку. В этом случае входящий Na+ нейтрализовал бы отрицательные заряды внутри клетки. Однако этого в действительности не происходит, так как мембрана в покое малопроницаема для Na+.
Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+, является натрий-калиевый насос (активный транспорт). Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается стремя находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с двумя находящимися вне клетки ионами К+ которые переносятся в цитоплазму. Энергообеспечение работы систем переносчиков обеспечивается АТФ. Функционирование насоса по такой системе приводит к следующим результатам:
- поддерживается высокая концентрация ионов К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя. Вследствие того что за один цикл обмена ионов из клетки выводится на один положительный ион больше, чем вводится, активный транспорт играет роль в создании потенциала покоя. В этом случае говорят об электрогенном насосе, поскольку он сам создает небольшой, но постоянный ток положительных зарядов из клетки, а потому вносит прямой вклад в формирование отрицательного потенциала внутри нее. Однако величина вклада электрогенного насоса в общее значение потенциала покоя обычно невелика и составляет несколько милливольт;
- поддерживается низкая концентрация ионов Na + внутри клетки, что, с одной стороны, обеспечивает работу механизма генерации потенциала действия, с другой — обеспечивает сохранение нормальных осмолярности и объема клетки;
- поддерживая стабильный концентрационный градиент Na + , натрий-калиевый насос способствует сопряженному К+, Na+ -транспорту аминокислот и Сахаров через клеточную мембрану.
Таким образом, возникновение трансмембранной разности потенциалов (потенциала покоя) обусловлено высокой проводимостью клеточной мембраны в состоянии покоя для ионов К + , СI-, ионной асимметрией концентраций ионов К + и ионов СI-, работой систем активного транспорта (Na+/K+ -АТФаза), которые создают и поддерживают ионную асимметрию.
Потенциал действия нервного волокна, нервный импульс
Потенциал действия - это кратковременное колебание разности потенциалов мембраны возбудимой клетки, сопровождающееся изменением ее знака заряда.
Потенциал действия является основным специфическим признаком возбуждения. Его регистрация свидетельствует о том, что клетка или ее структуры ответили на воздействие возбуждением. Однако, как уже отмечалось, ПД в некоторых клетках может возникать спонтанно (самопроизвольно). Такие клетки содержатся в водителях ритма сердца, стенках сосудов, нервной системе. ПД используется как носитель информации, передающий ее в виде электрических сигналов (электрическая сигнализации) по афферентным и эфферентным нервным волокнам, проводящей системе сердца, а также для инициирования сокращения мышечных клеток.
Рассмотрим причины и механизм генерации ПД в афферентных нервных волокнах, образующих первично воспринимающие сенсорные рецепторы. Непосредственной причиной возникновения (генерации) ПД в них является рецепторный потенциал.
Если измерять разность потенциалов на мембране ближайшего к нервному окончанию перехвата Ранвье, то в промежутках между воздействиями на капсулу тельца Пачини она остается неизменной (70 мВ), а во время воздействия деполяризуется почти одновременно с деполяризацией рецепторной мембраны нервного окончания.
При увеличении силы давления на тельце Пачини, вызывающей возрастание рецепторного потенциала до 10 мВ, в ближайшем перехвате Ранвье обычно регистрируется быстрое колебание мембранного потенциала, сопровождающееся перезарядкой мембраны — потенциал действия (ПД), или нервный импульс (рис. 2). Если сила давления на тельце возрастет еще больше, амплитуда рецепторного потенциала увеличивается и в нервном окончании генерируется уже ряд потенциалов действия с определенной частотой.

Рис. 2. Схематическое представление механизма преобразования рецепторного потенциала в потенциал действия (нервный импульс) и распространения импульса по нервному волокну
Суть механизма генерации ПД состоит в том, что рецепторный потенциал вызывает возникновение локальных круговых токов между деполяризованной рецепторной мембраной немиелинизированной части нервного окончания и мембраной первого перехвата Ранвье. Эти токи, носителями которых являются ионы Na+, К+, СI- и другие минеральные ионы, «протекают» не только вдоль, но и поперек мембраны нервного волокна в области перехвата Ранвье. В мембране перехватов Ранвье в отличие от рецепторной мембраны самого нервного окончания имеется большая плотность ионных потенциалзависимых натриевых и калиевых каналов.
При достижении на мембране перехвата Ранвье величины деполяризации около 10 мВ происходит открытие быстрых потенциалзависимых натриевых каналов и через них в аксоплазму по электрохимическому градиенту устремляется поток ионов Na+. Он обусловливает быструю деполяризацию и перезарядку мембраны перехвата Ранвье. Однако одновременно с открытием быстрых потенциалзависимых натриевых каналов в мембране перехвата Ранвье открываются медленные потенциалзависимые калиевые каналы и из аксоилазмы начинают выходить ионы К+ Их выход запаздывает по отношению ко входу ионов Na+. Таким образом, входящие с большой скоростью в аксоплазму ионы Na+ быстро деполяризуют и перезаряжают на короткое время (0,3-0,5 мс) мембрану, а выходящие ионы К+ восстанавливают исходное распределение зарядов на мембране (реполяризуют мембрану). В результате во время механического воздействия на тельце Пачини силой, равной или превышающей пороговую, на мембране ближайшего перехвата Ранвье наблюдается кратковременное колебание потенциала в виде быстрой деполяризации и реполяризации мембраны, т.е. генерируется ПД (нервный импульс).
Поскольку непосредственной причиной генерации ПД является рецепторный потенциал, то его в этом случае еще называют генераторным потенциалом. Число генерируемых в единицу времени одинаковых по амплитуде и длительности нервных импульсов пропорционально амплитуде рецепторного потенциала, а следовательно, силе давления на рецептор. Процесс преобразования информации о силе воздействия, заложенной в амплитуде рецепторного потенциала, в число дискретных нервных импульсов получил название дискретного кодирования информации.
Более подробно ионные механизмы и временная динамика процессов генерации ПД изучены в экспериментальных условиях при искусственном воздействии на нервное волокно электрическим током различной силы и длительности.
Природа потенциала действия нервного волокна (нервного импульса)
Мембрана нервного волокна в точке локализации раздражающего электрода отвечает на воздействие очень слабого тока, еще не достигшего порогового значения. Этот ответ получил название локального, а колебание разности потенциалов на мембране — локального потенциала.
Локальный ответ на мембране возбудимой клетки может предшествовать возникновению потенциала действия или возникать как самостоятельный процесс. Он представляет собой кратковременное колебание (деполяризация и реполяризация) потенциала покоя, не сопровождающееся перезарядкой мембраны. Деполяризация мембраны при развитиии локального потенциала обусловлена опережающим входом в аксоплазму ионов Na+, а реполяризация — запаздывающим выходом из аксоплазмы ионов К+.
Если воздействовать на мембрану электрическим током возрастающей силы, то при се величине, называемой пороговой, деполяризация мембраны может достигнуть критического уровня — Е к, при котором происходит открытие быстрых потенциалзависимых натриевых каналов. В результате через них происходит лавинообразно нарастающее поступление в клетку ионов Na+. Вызываемый процесс деполяризации приобретает самоускоряющийся характер, и локальный потенциал перерастает в потенциал действия.
Уже упоминалось, что характерным признаком ПД является кратковременная инверсия (перемена) знака заряда на мембране. Снаружи она на короткое время (0,3-2 мс) становится заряженной отрицательно, а внутри — положительно. Величина инверсии может составлять до 30 мВ, а величина всего потенциала действия — 60-130 мВ (рис. 3).
Таблица. Сравнительная характеристика локального потенциала и потенциала действия
|
Характеристика |
Локальный потенциал |
Потенциал действия |
|
Проводимость |
Распространяется местно, на 1-2 мм с затуханием (декрементом) |
Распространяется без затухания на большие расстояния по всей длине нервного волокна |
|
Закон «силы» |
Подчиняется |
Не подчиняется |
|
Закон «все или ничего» |
Не подчиняется |
Подчиняется |
|
Явление суммации |
Суммируется, возрастает при повторных частых подпороговых раздражениях |
Не суммируется |
|
Величина амплитуды |
||
|
Способность к возбудимости |
Увеличивается |
Уменьшается вплоть до полной невозбудимости (рефрактерность) |
|
Величина раздражителя |
Подпороговая |
Пороговая и сверхпороговая |
Потенциал действия в зависимости от характера изменения зарядов на внутренней поверхности мембраны подразделяют на фазы деполяризации, реполяризации и гиперполяризации мембраны. Деполяризацией называют всю восходящую часть ПД, на которой выделяют участки, соответствующие локальному потенциалу (от уровня Е 0 до Е к ), быстрой деполяризации (от уровня Е к до уровня 0 мВ), инверсии знака заряда (от 0 мВ до пикового значения или начала реполяризации). Реполяризацией называют нисходящую часть ПД, которая отражает процесс восстановления исходной поляризации мембраны. Вначале реполяризация осуществляется быстро, но, приближаясь к уровню Е 0 , скорость се может замедляться и этот участок называют следовой отрицательностью (или следовым отрицательным потенциалом). У некоторых клеток вслед за реполяризацией развивается гиперполяризация (возрастание поляризации мембраны). Ее называют следовым положительным потенциалом.
Начальную высокоамплитудную быстропротекающую часть ПД называют также пик, или спайк. Он включает фазы деполяризации и быстрой реполяризации.
В механизме развития ПД важнейшая роль принадлежит потенциалзависимым ионным каналам и неодновременному увеличению проницаемости клеточной мембраны для ионов Na+ и К+. Так, при действии на клетку электрического тока он вызывает деполяризацию мембраны и, когда заряд мембраны уменьшается до критического уровня (Е к), открываются потенциалзависимые натриевые каналы. Как уже упоминалось,эти каналы образованы встроенными в мембрану белковыми молекулами, внутри которых имеются пора и два воротных механизма. Один из воротных механизмов — активационный обеспечивает (при участии сегмента 4) открытие (активацию) канала при деполяризации мембраны, а второй (при участии внутриклеточной петли между 3-м и 4-м доменами) — его инактивацию, развивающуюся при перезарядке мембраны (рис. 4). Поскольку оба этих механизма быстро изменяют положение ворот канала, то потенциалзависимые натриевые каналы являются быстрыми ионными каналами. Это обстоятельство имеет определяющее значение для генерации ПД в возбудимых тканях и для его проведения по мембранам нервных и мышечных волокон.

Рис. 3. Потенциал действия, его фазы и ионные токи (а, о). Описание в тексте

Рис. 4. Положение ворот и состояние активности потенциалзависимых натриевого и калиевого каналов при различных уровнях поляризации мембраны
Чтобы потенциалзависимый натриевый канал мог пропускать внутрь клетки ионы Na+, необходимо открыть лишь активационные ворота, поскольку инактивационные в условиях покоя открыты. Это и происходит, когда деполяризация мембраны достигает уровня Е к (рис. 3, 4).
Открытие активационных ворот натриевых каналов приводит к лавинообразному вхождению натрия внутрь клетки, движимому действием сил его электрохимического градиента. Поскольку ионы Na+ несут положительный заряд, то они нейтрализуют избыток отрицательных зарядов на внутренней поверхности мембраны, снижают разность потенциалов на мембране и деполяризуют ее. Вскоре ионы Na+ придают внутренней поверхности мембраны избыток положительных зарядов, что сопровождается инверсией (сменой) знака заряда с отрицательного на положительный.
Однако натриевые каналы остаются открытыми лишь около 0,5 мс и через этот промежуток времени от момента начала
ПД закрываются инактивационные ворота, натриевые каналы становятся инактивированными и непроницаемыми для ионов Na+, поступление которых внутрь клетки резко ограничивается.
С момента деполяризации мембраны до уровня Е к наблюдаются также активация калиевых каналов и открытие их ворот для ионов К+. Ионы К+ под действием сил концентрационного градиента выходят из клетки, вынося из нее положительные заряды. Однако воротный механизм калиевых каналов является медленно функционирующим и скорость выхода положительных зарядов с ионами К+ из клетки наружу запаздывает по отношению ко входу ионов Na+. Поток ионов К+, удаляя из клетки избыток положительных зарядов, обусловливает восстановление на мембране исходного распределения зарядов или ее реполяризацию, и на се внутренней стороне через мгновение от момента перезарядки восстанавливается отрицательный заряд.
Возникновение ПД на возбудимых мембранах и последующее восстановление исходного потенциала покоя на мембране оказываются возможными потому, что динамика входа в клетку и выхода из клетки положительных зарядов ионов Na+ и К+ различна. Вход иона Na+ по времени опережает выход иона К+. Если бы эти процессы были равновесными, то разность потенциалов на мембране не изменялась бы. Развитие способности к возбуждению и генерации ПД возбудимыми мышечными и нервными клетками было обусловлено формированием в их мембране двух типов разноскоростных ионных каналов — быстрых натриевых и медленных калиевых.
Для генерации одиночного ПД требуется поступление в клетку относительно небольшого числа ионов Na+, которое не нарушает его распределения вне и внутри клетки. При генерации большого числа ПД распределение ионов по обе стороны мембраны клетки могло бы нарушиться. Однако в нормальных условиях это предотвращается работой Na+, К+ -насоса.
В естественных условиях в нейронах ЦНС потенциал действия первично возникает в области аксонного холмика, в афферентных нейронах — в ближайшем к сенсорному рецептору перехвате Ранвье нервного окончания, т.е. в тех участках мембраны, где имеются быстрые селективные потенциалзависимые натриевые каналы и медленные калиевые каналы. В других типах клеток (например, пейсмекерных, гладких миоцитах) в возникновении ПД играют роль не только натриевые и калиевые, но и кальциевые каналы.
Механизмы восприятия и преобразования в ПД сигналов во вторично чувствующих сенсорных рецепторах отличаются от механизмов, разобранных для первично чувствствующих рецепторов. В этих рецепторах восприятие сигналов осуществляется специализированными нейросенсорными (фоторецепторные, обонятельные) или сенсоэпителиальными (вкусовые, слуховые, вестибулярные) клетками. В каждой из этих чувствительных клеток имеется свой, особый механизм восприятия сигналов. Однако во всех клетках энергия воспринимаемого сигнала (раздражителя) преобразуется в колебание разности потенциалов плазматической мембраны, т.е. в рецепторный потенциал.
Таким образом, ключевым моментом в механизмах преобразования сенсорными клетками воспринимаемых сигналов в рецепторный потенциал является изменение проницаемости ионных каналов в ответ на воздействие. Открытие Na+, Са 2+ , К+ -ионных каналов при восприятии и преобразовании сигнала достигается в этих клетках при участии G-белков, вторых внутриклеточных посредников, связывании с лигандами, фосфорилировании ионных каналов. Как правило, возникший в сенсорных клетках рецепторный потенциал вызывает высвобождение из них в синаптическую щель нейромедиатора, который обеспечивает передачу сигнала на постсинаптическую мембрану афферентного нервного окончания и генерацию на его мембране нервного импульса. Эти процессы подробно описаны в главе, посвященной сенсорным системам.
Потенциал действия может быть охарактеризован амплитудой и продолжительностью, которые для одного и того же нервного волокна остаются одинаковыми при распространении ПД по волокну. Поэтому потенциал действия называют дискретным потенциалом.
Между характером воздействия на сенсорные рецепторы и числом ПД, возникших в афферентном нервном волокне в ответ на воздействие, имеется определенная связь. Она заключается в том, что на большие но силе или продолжительности воздействия в нервном волокне формируется большее число нервных импульсов, т.е. при усилении воздействия в нервную систему будут посылаться от рецептора импульсы большей частоты. Процессы преобразования информации о характере воздействия в частоту и другие параметры нервных импульсов, передаваемых в ЦНС, получили название дискретного кодирования информации.
1. Барьерная (защищает клетку, поддерживает ее форму)
2. Транспортная (определяет состав веществ внутри клетки)
3. Рецепторная (определяет специальную чувствительность данной клетки к определенной группе химических веществ)
4. Электрическая (обеспечивает создание разности потенциалов между внутренней и внешней поверхностью мембраны).
Отличительные свойства нервной клетки:
Возбудимость (способность генерировать потенциал действия при раздражении)
Проводимость (способность проводить и передавать возбуждение другим клеткам).
Электрические процессы в нейронах
Природа мембранного потенциала (потенциала покоя)
Потенциал покоя формируется благодаря пассивному (по градиентам) выходу ионов калия из клетки. В результате: -на наружной поверхности мембраны возникает избыток положительно заряженных ионов; внутри клетки остаются отрицательно заряженные крупные молекулы.
Механизм возбуждения нейрона:
1. Начальное изменение потенциала мембраны;
2. раскрывается часть натриевых каналов;
3. повышается проницаемость мембраны для натрия;
4. перемещение натрия в клетку по электрическому и химическому градиентам.
5. Рост числа положительны ионов внутри клетки;
6. Локальная деполяризация мембраны. (если деполяризация незначительна, то все сначала)
Если деполяризация достигает критической величины:
7. Раскрываются все натриевые каналы;
8. Происходит резкая деполяризация мембраны - потенциал действия (от -90 мВ до +30 мВ)
9. Натриевые каналы захлопываются и раскрываются калиевые каналы (через 0,5 мс).
10. Прекращается диффузия натрия, и начинается выход калия, который вытягивает электрический градиент.
11. Восстановление мембранного потенциала до исходных значений – реполяризация.
12. Прекращение выхода калия из клетки за счет изменения электрического градиента.
13. Включается натрий-калиевый насос.
14. Восстановление исходного ионного баланса (калий – внутри клетки, натрий – снаружи).
Ионный насос – мембранная транспортная система, обеспечивающая перенос ионов против электрохимического градиента, то есть с затратой энергии.
[рис. Потенциал действия]
Закон «все или ничего»
Если деполяризация мембраны достигает критической (пороговой) величины, то формируется потенциал действия. Если деполяризаця мембраны не достигает пороговой величины, то потенциал действия не формируется.
Преимущества сальтаторного проведения:
Экономичность (площадь перехвата менее 1% мембраны аксона)
Скорость (поле распространяется на большее расстояние).
Структурные элементы синапса:
1. Пресинаптическая мембрана (мембрана аксона, передающего нейрона)
2. Синаптическая щель (межклеточная жидкость)
3. Постсинаптическая мембрана (мембрана дендрита или сомы принимающего нейрона)
Механизм синаптической передачи.
1. Приход потенциала действия в синаптическое окончание аксона;
2. Раскрытие кальциевых каналов;
3. Повышение проницаемости мембраны для кальция;
4. Перемещение ионов кальция в клетку;
5. Деполяризация пресинаптической мембраны;
6. Выброс медиаторов в синаптическую щель (чем больше деполяризация – тем больше выброс).
7. Соединение медиатора со специфическими рецепторами постсинаптической мембраны;
8. Изменение потенциала постсинаптической мембраны;
9. Раскрытие ионных каналов;
10. Если увеличение проницаемости ионов натрия приводит к формированию ВПСП (возбудительный постинаптический потенциал), если калия и хлора – ТПСП.
Проведение в синапсах: одностороннее, с задержкой.
Свойства постсинаптических потенциалов:
Градуальность (амплитуда потенциалов переменная и отражает частоту потенциалов действия, поступающих на синапс)
Локальность (ВПСП и ТПСП распространяются по нейрону с затуханием).
Способность к суммации (суммируются потенциалы, близкорасположенные в пространстве и времени).
Функции нейроглии:
Защитная
Изолирующая
Обменная.
Функции глиальных клеток:
1. Астроциты: формируют каркас для нейронов; обеспечивают метаболизм; регенерация нерва.
2. Олигодендроциты: миелиновые оболочки аксонов.
Функционирование спинного мозга
Спинной мозг – это главный исполнительный отдел ЦНС. В его задачи входит передача команд на мышцы и железы, а также регуляция работы внутренних органов.
Корешки спинного мозга делятся на задние и передние.
Задние – чувствительные– афферентные. Состоят из аксонов клеток спинальных ганглиев. По ним предается информация от кожных рецепторов, проприорецепторов, висцерорецепторов.
Передние – двигательные – эфферентные. Состоят из аксоны мотонейронов. Направляются к мышцам к железам.
Каждый сегмент спинного мозга иннервирует три метамера тела.
Задние рога спинного мозга состоят из чувствительных (афферентых) нейронов, интернейронов (вставочных нейронв), а также клеток желатинозной субстанции (тормозные нейроны).
Передние рога состоят из мотонейронов.
Функциональные отделы серого вещества спинного мозга [рисунок]
По восходящим путям спинного мозга передаются:
- сигналы от рецепторов мышц и сухожилий (проприорецепторы) по пучкам Голяя и Бурдаха, по спиномозжечковым путям Говерса и Флексига.
Сигналы от болевых и тепературных рецепторов по латеральному спиноталамическому тракту.
Сигналы от тактильных рецепторов по вентральному спиноталамическому пути и частичто по пучкам Голля и Бурдаха.
Нисходящие пути спинного мозга. Включают в себя две системы: пирамидную и экстарпирамидную систему.
По пирамидной системе передается команды на выполнение целенаправленных движений, по кортикоспинальным путям.
По экстарпирамидной системе передаются команды поддержания позы и равновесия по ретикулоспинальным, руброспинальным, тектоспинальным, вестибулоспинальным и оливоспинальным путям.
Спинной мозг реализует две основных функции: рефлекторная и проводниковая.
Рефлекторная функция спинного мозга
Рефлекс – это стереотипная реакция организма на раздражение рецепторов, осуществляемая при участии нервной системы.
Дуга спинального соматического рефлекса [рисунок]
Фунзкциональной единицей спинного мозга является цепь, объединяющая чувствительный нейрон с мотонейроном.
Рефлекторная дейятельность спинного мозга обеспечиваеется :
1. передачей возбуждения с чувствительных нейронов на моторные нейроны.
2. Регуляцией передачи возбуждения в рефлекторной дуге.
Регуляция рефлексов осуществляется через организованное торможения.
1. Внутрисементарное торможение – скоординированная работа флексоров и экстензоров (сгибатели и разгибатели).
2. Межсегментарное тороможение – скоординировання работа мыщечных групп.
3. Эфферентное (центральное) торможение внутриспинальных тормозных связей – сила и скорость рефлекторной реакции.
Реализация рефлекторной функции:
1. обработка афферентных сигналов
2. обработка команд от управляющих структур
3. Формирование моторных команд
4. Формирование обратной афферентации
Рефлексы спинного мозга:
1. Мышечные (движения, поза).
2. Кожные (сосудистые, потоотделительные…)
3. Висцеральные (мышцы груди, спины…)
Спинальные рефлексы:
1. Простые (реализуются одним сегментом)
2. Сложные (реализуются несколькими сегментами)
Спинной мозг – это центр элементарных двигательных программ; центр врожденных двигательных автоматизмов;
Проводниковая функция спинного мозга
Заключается в передаче восходящих и нисходящих потоков информации. Восходящая информация – о положении конечностей, туловища, головы. Нисходящая информация содержит в себе команды на движения, поддержание позы и равновесия, регулирующие влияния (на рефлекторные дуги).
Вегетативная нервная система спинного мозга
Вегетативная нервная система – это та часть нервной системы, которая иннервирует внутренние органы, сосуды, железы и мышцы.
Состоит из двух отделов:
Симпатической системы (локализована в спинном мозге - в боговых рогах)
Парасимпатической системы (локализована в стволе головного мозга и в крестцовом отделе спинного мозга – в боковых рогах)
Центры симпатической НС
1. Грудной отдел спинного мозга (боковые рога)
2. Поясничные отдел спинного мозга (боковые рога)
Центры парасимпатической НС, находящиеся в спинном мозге, находятся в крестцовом отделе спинного мозга (боковые рога).